



【プレスリリース】メラニン合成酵素の分解を促す新分子を発見~新分子をターゲットにした新たな美白剤への応用に期待~
ポイント
▪ メラニン合成酵素のメラノソームへの輸送を制御する Varp 分子の新たな結合パートナーとして Rab40C を同定
▪ Rab40C は Varp 分子のユビキチン化を促し、プロテアソームでの分解を促進
▪ Rab40C の発現を増加させたメラノサイトでは、Varp 分子の分解が促進されるため、メラニン合成酵素の輸送が阻害され、結果的にメラニン合成酵素の分解も促進
概要
国立大学法人東北大学は、メラニン合成酵素の分解を促す新分子を同定することに成功しました。これは、東北大学大学院生命科学研究科の谷津彩香修士大学院生、島田光修士大学院生、大林典彦助教、福田光則教授による研究成果です。
有害な紫外線からわたしたちの体を守るメラニン色素は、メラノソーム*1 と呼ばれる特殊な袋(小胞)の中でメラニン合成酵素によって合成されています。メラニン合成酵素はメラノソームにはじめから存在する訳ではなく、メラノソームの袋の元ができてから、後からメラノソームへ輸送されることが知られています。私達はこれまで、この輸送を制御する分子としてVarp(バープ)*2 を同定しており、この分子の発現異常によりメラニン合成酵素の分解(すなわち、メラニン色素量の減少)が促進されることを見出しています。かし、メラノサイトにおけるVarp 分子の発現調節に関わる仕組みはこれまで全く分かっていませんでした。
今回、研究グループはVarp に結合する新たな分子として、Rab40C*3 を同定することに成功しました。Rab40C はVarp 分子のユビキチン化を促進し、プロテアソームと呼ばれるタンパク質分解酵素*4 でVarp 分子を分解に導くことが明らかになりました。したがって、Rab40C をメラノサイトに過剰に発現させるとVarp 分子の分解が、逆にRab40C の発現を低下させるとVarp 分子の蓄積が観察されますが、メラニン合成酵素の輸送には適切な量のVarp が必要なため、いずれの場合でもメラニン合成酵素がメラノソームに正しく輸送されず、結果的に分解されてしまうことを突き止めました。
今回の『メラニン合成酵素には直接作用せず、メラニン合成酵素の分解を促進する分子』の発見により、従来の「メラニン合成酵素の直接的な酵素活性の阻害」とは異なるアプローチの美白化粧品の開発に応用されることが期待されます。
本研究成果は、英国の科学雑誌『Biology Open』電子版に間もなく掲載される予定です。
背景
わたしたちの肌や髪の毛の色の源であるメラニン色素は、有害な紫外線からわたしたちの体を守るために重要な役割を果たしています。しかし、その一方でメラニン色素の肌への過剰な沈着はしみやそばかすの原因にもなっています。メラニン色素は、メラノサイト(メラニン色素産生細胞)に存在する「メラノソーム」*1 と呼ばれる袋(小胞)の中でメラニン合成酵素によって合成されています。メラニン合成酵素はメラノソームの袋にはじめから含まれている訳ではなく、メラノソームとは別個に作られ、後からメラノソーム内に運び込まれています(図1)。この運搬過程には、「Varp(バープ)」*2 と呼ばれる分子が関与しており、この分子の発現異常(発現低下あるいは過剰な発現上昇)によりメラニン合成酵素の分解(すなわち、メラニン色素量の減少)が促進されることが明らかになっています。しかし、メラノサイトにおけるVarp 分子の発現調節(特に分解制御)に関わる仕組みはこれまで全く分かっていませんで した。
研究成果
これまでの研究で、Varp 分子には4つの機能ドメイン(アミノ末端側からVPS9 ドメイン、ANKR1 ドメイン、VID ドメイン、ANKR2 ドメイン)が存在し、アミノ末端側の3つのドメインを介してメラノサイトの樹状突起の形成やメラニン合成酵素の輸送を制御することが明らかになっています(図2)*2。一方、Varp 分子のカルボキシル末端側に存在するANKR2 ドメインの機能はこれまで全く分かっていませんでした。本研究ではまず、ANKR2 ドメインに結合する分子の探索を行い、低分子量G タンパク質の一種Rab40*3 が特異的に結合することを見出しました(図3A)。構造上、Rab40 はSOCS ボックス*3 と呼ばれるタンパク質分解に関わるユビキチンリガーゼ複合体*4 を結合する領域を持っていたことから、次にVarp 分子の分解に対する影響を検討しました(図4~図7)。その結果、以下のことを明らかにすることができました。
1. マウス培養メラノサイトにはRab40C が内在性に発現しており、活性化型(GTP 結合型)依存的にVarp 分子と結合します(図3B,C)。
2. Rab40C の野生型あるいは常時活性化型変異体(Q73L)をマウス培養メラノサイトに発現させると、Varp 分子の発現量が顕著に低下します(図4、矢印の細胞)。一方、Varp との結合能をほとんど示さない常時不活性化型変異体(G28N)やSOCS ボックスを欠損させたΔSOCS 変異体(Varp との結合能は正常)では、Varp の発現量には特に影響は見られません。
3. Rab40C はVarp 分子のポリユビキチン化を促進し、プロテアソーム(タンパク質分解酵素)での分解を誘導します。このため、Rab40C を発現するメラノサイトをプロテアソームの阻害剤で処理すると、Varp 分子の分解が抑制されます(図5)。
4. Rab40C の野生型あるいは常時活性化型変異体を発現するメラノサイトでは、Varp の発現量が低下しているため、Varp によってメラノソームに輸送されるメラニン合成酵素の一種チロシナーゼ関連タンパク質1(Tyrp1)の発現量も顕著に低下します(図6、矢印の細胞)。
5. メラノサイトに内在性のRab40C をRNA 干渉法によりノックダウンすると(図7A)、Varp分子の分解が抑制されるため、その発現量が増加します(図7B)。メラニン合成酵素の輸送には適切な量のVarp 分子が必要なため、過剰なVarp 分子を発現するRab40C 欠損メラノサイトでは、メラニン合成酵素がメラノソーム上から消失してしまいます(図7C、矢印の細胞)。
以上の結果から、Rab40C はVarp 分子のメラノサイトでの発現量を厳密に制御する役割を担うものと考えられました。Varp 分子はメラニン合成酵素のメラノソームへの輸送に必須の役割を果たすため、Rab40C の発現異常(過剰発現あるいは発現低下)によりVarp 分子の発現量が適切に保てなくなり、間接的にメラニン合成酵素のメラノソームでの発現に影響を与えることが明らかになりました。
今後の展開
今回の研究により、メラノサイトにはVarp 分子を分解することによりメラニン合成酵素の輸送を抑制する機構が備わっていることが明らかになりました。これまでの美白化粧品の多くは「メラニン合成酵素の基質に類似した化合物による直接的な酵素活性の阻害」という観点から作製されており、一部の化合物ではメラニン合成酵素により有害な物質に代謝される危険性が指摘されていました。今回の『メラニン合成酵素には直接作用せず、メラニン合成酵素の分解を促進するRab40C 分子』の発見により、従来のアプローチとは異なる美白化粧品の開発が可能と考えられます。今後、Rab40C などVarp 分子の分解に関与するタンパク質をターゲットとした薬剤スクリーニングが進めば、メラニン合成酵素の分解を促し、しみ・そばかすの予防、美白の維持が可能になるものと期待されます。
※本研究成果は、公益財団法人東京生化学研究会・研究助成金「メラノソームダイナミクスを制御する分子基盤の解明と創薬へのアプローチ」(研究代表者:福田光則 東北大学大学院生命科学研究科教授)、公益財団法人 東レ科学技術振興会・研究助成金「リソソーム関連オルガネラ輸送の分子機構の解明」(研究代表者:福田光則 東北大学大学院生命科学研究科教授)、独立行政法人日本学術振興会・学術研究助成基金助成金 基盤研究C「色素細胞活性化におけるVarp の機能解析」(研究代表者:大林典彦 東北大学大学院生命科学研究科助教)のサポートによるものです。
□ PDF
□ 東北大学
□ 生命科学研究科
□ 生命科学研究科 膜輸送機構解析分野
図及び説明
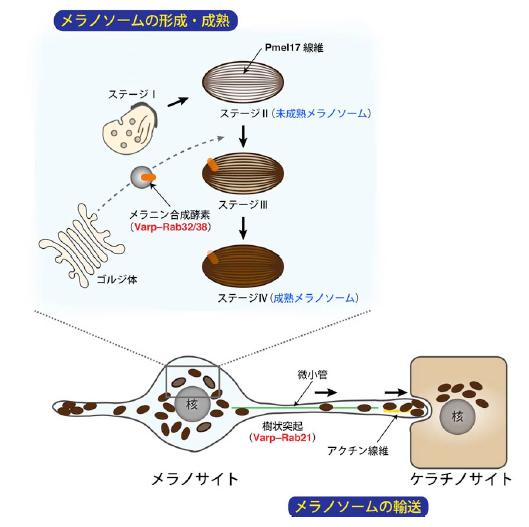 図1 メラノサイトにおけるメラノソームの形成・成熟・輸送の仕組み
図1 メラノサイトにおけるメラノソームの形成・成熟・輸送の仕組み
メラニン色素を合成・貯蔵するメラノソームは、その成熟度合いによって形態学的に4つのステージに分類されています。ステージⅠ、Ⅱの段階(未成熟メラノソーム)では、メラノソームの元となる袋とメラニン色素が将来沈着するPmel17 線維が形成されていますが、メラニン合成酵素が無いため色は付いていません。メラニン合成酵素は未成熟メラノソームとは別個に合成され、Varp-Rab32/38(*2 参照)の作用により未成熟メラノソームへと輸送されます。その後、メラニン色素がPmel17 線維に沈着して行き(ステージⅢ)、やがて黒いメラノソームへと成熟します(ステージⅣ)。核周辺で成熟したメラノソームは、細胞内の二種類の道路(細胞骨格:微小管とアクチン線維)に沿って細胞膜の直下まで輸送され、最終的に隣接するケラチノサイト(あるいは毛母細胞)に受け渡されて肌(あるいは髪の毛)の暗色化が起こります。なお、微小管上の輸送は順行性と逆行性の両方向性に輸送されますが、アクチン線維上の輸送は一方向に起こります。これまでの本研究グループの解析で、微小管順行性輸送にはRab1Aが、微小管逆行性輸送にはメラノレギュリンやRab36 が、またアクチン輸送にはRab27A が関与することが既に明らかになっています。
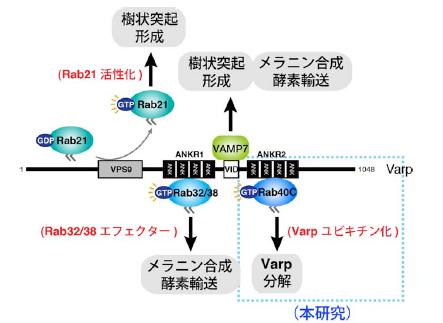 図2 Varp 分子の構造と機能
図2 Varp 分子の構造と機能
マウスのVarp 分子は1048 アミノ酸からなり、アミノ末端側からRab21 を活性化するVPS9 ドメイン、活性化型のRab32/38 を結合するANKR1 ドメイン、VAMP7 を結合するVID ドメイン、及びこれまで機能未知のANKR2 ドメインにより構成されています(*2 参照)。これまでの研究で、Varp 分子はメラノサイトの樹状突起の形成とメラニン合成酵素(チロシナーゼやTyrp1)の輸送に関与することが明らかになっています。本研究により、ANKR2 ドメインにはRab40C(*3 参照)が結合することにより、Varp 分子の分解を制御することが明らかになっています。
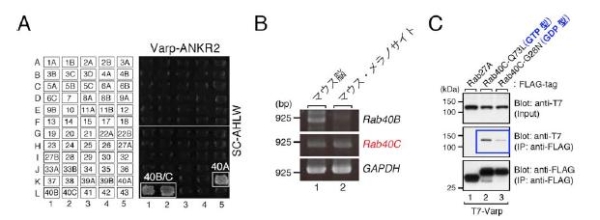 図3 Varp ANKR2 ドメインの新規結合分子Rab40 の同定
図3 Varp ANKR2 ドメインの新規結合分子Rab40 の同定
(A)Varp 分子のANKR2 ドメインはRab32/38 を結合するANKR1 ドメインとも相同性があり、酵母two-hybrid スクリーニングの結果、ANKR2 ドメインにはRab40 のサブファミリー(Rab40A/B/C)が特異的に結合することが明らかになりました。(B)Rab40A はマウスには保存されていないため、Rab40B/C のマウスメラノサイトにおける発現をRT-PCR 法により確認したところ、Rab40C が主に発現していました。(C)Rab40C とVarp の結合はGTP 依存的で、常時不活性化型変異体(G28N)よりも常時活性化型変異体(Q73L)に強く結合しました。
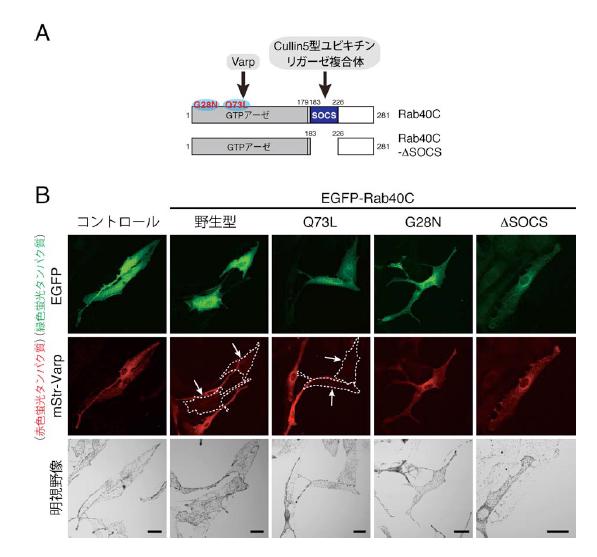 図4 Rab40C の構造とVarp の発現への影響
図4 Rab40C の構造とVarp の発現への影響
(A)Rab40C を含むRab40 のサブファミリーは他のRab とは異なり、GTP アーゼドメイン(GTP/GDP を結合する領域)に加え、SOCS ボックスと呼ばれるCullin5 型のユビキチンリガーゼ複合体を結合する領域を有しています(*3 及び*4 参照)。Varp はRab40C のGTP アーゼドメインに結合しますので、SOCS ボックスを欠損させたΔSOCS 変異体でも野生型と同じようにVarp との結合が観察されます。28 番目のグリシン(G)をアスパラギン(N)に置換したG28N変異体は常時不活性化型(GDP 結合型)として、73 番目のグルタミン(Q)をロイシン(L)に置換したQ73L 変異体は常時活性化型(GTP 結合型)として作用します。(B)マウス培養メラノサイト(melan-a 細胞)に緑色蛍光タンパク質(EGFP)を付加したRab40C(野生型及び各種変異体)と赤色蛍光タンパク質(mStr)を付加したVarp を発現させ、Varp の発現に対する影響を蛍光観察により検討しました。Varp との結合能を有する野生型Rab40C やQ73L 変異体を発現する細胞では、Varp の発現が顕著に低下していることが明らかになりました(点線の細胞、矢印)。一方、Varp との結合能が弱いG28N 変異体やCullin5 型ユビキチンリガーゼ複合体と結合が出来ないΔSOCS 変異体では、このようなVarp の発現低下は観察されませんでした。スケールバー = 20μm。
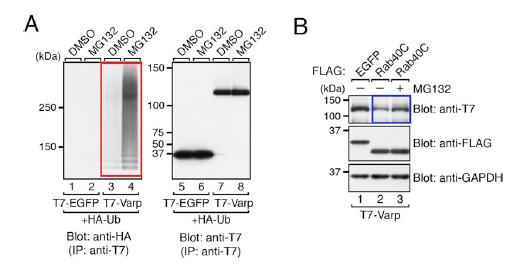 図5 Varp 分子のユビキチン化とRab40C 依存的な分解
図5 Varp 分子のユビキチン化とRab40C 依存的な分解
(A)メラノサイトに発現したVarp 分子のポリユビキチン化(赤枠)。プロテアソーム(*4 参照)の阻害剤であるMG132 で処理すると、Varp 分子のポリユビキチン化(HA タグ抗体で検出)がより顕著になります(レーン3 と4 の比較)。(B)Varp 分子のメラノサイトでの発現量はRab40C の発現により低下し、この低下はMG132 処理により相殺されます(青枠)。
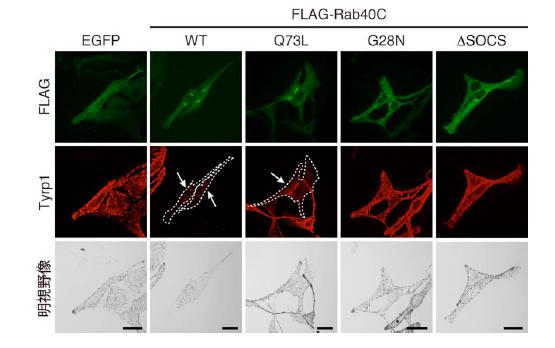 図6 Rab40C 発現によるメラニン合成酵素Tyrp1 の発現低下
図6 Rab40C 発現によるメラニン合成酵素Tyrp1 の発現低下
マウス培養メラノサイトに緑色蛍光タンパク質(EGFP)を付加したRab40C(野生型及び各種変異体)を発現させ、メラニン合成酵素Tyrp1 の発現に対する影響を免疫染色法により検討しました。野生型Rab40C やQ73L 変異体を発現する細胞では、Varp 分子の発現が低下するため、Tyrp1 の発現が顕著に低下していることが明らかになりました(点線の細胞、矢印)。一方、G28N 変異体やΔSOCS 変異体では、Varp の発現量に影響を与えないため、このようなTyrp1 の発現低下は観察されませんでした。スケールバー = 20μm。
 図7 Rab40C 欠損細胞におけるメラニン合成酵素Tyrp1 の発現低下
図7 Rab40C 欠損細胞におけるメラニン合成酵素Tyrp1 の発現低下
(A)RNA 干渉法を用いメラノサイトに内在性のRab40C のmRNA をノックダウンしても(赤枠)、Varp のmRNA の発現量には影響はありません。(B)Rab40C を欠損させたメラノサイトでは、Varp 分子の分解が抑制されるため、Varp 分子のタンパク質発現量が増加します(青枠)。メラニン合成酵素のメラノソームへの輸送には適切な量のVarp が必要なため、Varp 分子が過剰に発現した状態ではメラニン合成酵素Tyrp1 を正しくメラノソームに輸送できず、細胞内で分解されてしまいます。(C)このため、Rab40C を欠損させたメラノサイトではTyrp1 のシグナルの顕著な低下が観察されます(点線の細胞、矢印)。スケールバー = 20μm。
用語説明
*1 メラノソーム
細胞内には膜で包まれた細胞内小器官(オルガネラ)と呼ばれる袋状の構造物が多数存在しています。メラノソームはメラニン色素を合成・貯蔵するのに特化したオルガネラで、メラノサイト(メラニン色素産生細胞)と呼ばれる特殊な細胞にのみ存在しています。メラニン色素を合成するメラニン合成酵素(チロシナーゼやチロシナーゼ関連タンパク質1(Tyrp1)など)は、はじめからメラノソームに存在する訳ではなく、メラノソームの袋の元(未成熟メラノソーム)ができてから、後からメラノソームへ輸送されます。成熟した黒いメラノソームは、その後メラノサイトの細胞内を細胞骨格に沿って輸送され(図1)、肌や髪の毛を作る細胞に受け渡され、はじめて肌や髪の毛の暗色化が起こります。
*2 Varp(バープ)
Varp(VPS9-ankyrin-repeat protein)は分子内にVPS9 ドメインと呼ばれるRab21(*3 参照)の活性化ドメイン(グアニンヌクレオチド交換因子)、2個のアンキリンリピートドメイン(様々なタンパク質に見られる、33 アミノ酸の繰り返し配列で、分子間・分子内の相互作用に寄与)、VAMP7 を結合するVID ドメインを持つタンパク質です(図2)。これまでの研究で、Varp はVPS9 ドメインで Rab21 の活性化を行い樹状突起の形成を促進し、最初のアンキリンリピート(ANKR1)ドメインで活性化型のRab32/38 を結合し(Rab32/38 のエフェクター分子として機能)、メラニン合成酵素のメラノソームへの輸送を制御すること、VID ドメインは樹状突起の形成とメラニン合成酵素の輸送の両方のプロセスに関与することが明らかになっています。一方、カルボキシル末端側のANKR2 ドメインの機能はこれまで明らかではありませんでした。本研究により、ANKR2 ドメインにはRab40C(*3 参照)が結合し、Varp 分子のプロテアソームでの分解(*4 参照)を制御することが初めて明らかになりました。
プレスリリース(2009年4月28日)
「メラニン合成酵素」を輸送する新分子発見 --新たな美白ターゲットとして期待--
http://www.tohoku.ac.jp/japanese/newimg/pressimg/20090428.pdf
プレスリリース(2011年12月13日)
Varp 分子の新たな機能「樹状突起形成の促進作用」--メラニン色素のケラチノサイトへの転移に関与--
http://www.tohoku.ac.jp/japanese/newimg/pressimg/%28HP%29tohokuuniv-press20111213_2.pdf
プレスリリース(2013年4月24日)
メラニン合成酵素をメラノソームに受け渡す仕組み解明へ ~メラニン合成酵素の受け渡しに関わる膜融合装置SNARE~
http://www.tohoku.ac.jp/japanese/newimg/pressimg/tohokuuniv-press_20130422_01web.pdf
*3 低分子量G タンパク質Rab40C とSOCS ボックス
膜で包まれた小胞(メラノソームなど)の輸送を適切に行うためには交通整理人(制御タンパク質)の存在が不可欠です。この交通整理人の一つとして酵母から高等哺乳動物のヒトまで普遍的に存在しているのが、低分子量G タンパク質Rab(ラブ)です。Rab はGTP というヌクレオチドを結合した活性化型とGDP を結合した不活性化型の二つの状態をとり、活性化型の GTP-Rab(例えばGTP-Rab32/38)がエフェクターと呼ばれる特異的なタンパク質(例えばVarp ANKR1)と結合することにより小胞(例えばメラニン合成酵素を含む小胞)を輸送します。不活性化型のGDP-Rab をGTP-Rab に変換し、活性化を行う酵素をグアニンヌクレオチド交換因子(GEF)と呼び、Varp のVPS9 ドメインはRab21 の活性化を行います。Rab40C もRab の一種ですが、他のRab とは異なり、SOCS(suppressor of cytokine signaling)ボックスと呼ばれるCullin5 型のユビキチンリガーゼ複合体(*4 参照)と結合する領域をさらに持っています(図4A)。しかし、哺乳動物のRab40 のSOCS ボックスの機能はこれまで明らかではありませんでした。今回の研究により、Rab40C はCullin5 型ユビキチンリガーゼ複合体と基質となるVarp の両方に結合し、Varp のユビキチン化と分解を促進することが明らかになりました。
*4 ユビキチン-プロテアソーム
2004年のノーベル化学賞の対象となった生命現象で、ユビキチンリガーゼ(今回の場合、Cullin5 型ユビキチンリガーゼ複合体)の作用により、ユビキチン(76 アミノ酸からなるタンパク質)が標的タンパク質(今回の場合、Varp)のリジン残基に付加されます。ユビキチンのリジン残基にはさらに次のユビキチンが付加され、ポリユビキチン化された標的タンパク質はプロテアソームと呼ばれるタンパク質分解酵素で分解されます。
論文題目
Yatsu, A., Shimada, H., Ohbayashi, N. & Fukuda, M. (2015) Rab40C is a novel Varp-binding protein that promotes proteasomal degradation of Varp in melanocytes. Biol. Open, in press (DOI:10.1242/bio.201411114)
「メラノサイトにおいてRab40C はVarp 分子に結合し、プロテアソームでの分解を促進する」
問い合わせ先
東北大学大学院生命科学研究科 教授 福田 光則 (ふくだ みつのり)
Email: nori[at]m.tohoku.ac.jp TEL: 022-795-7731(or 022-795-3641)
東北大学大学院生命科学研究科広報室 広報担当 高橋 さやか
Email: lifsci-pr[at]ige.tohoku.ac.jp TEL: 03-5734-2975 FAX: 03-5734-3661
※[at]を「@」に置き換えてください。
Posted on:2015年2月 5日