
お知らせ

- トップ
- お知らせ
アフリカの栽培イネが芒(のぎ)を失った理由
~アジアとアフリカで異なる遺伝子の選抜が起きたことを解明~
発表のポイント
● イネの芒(のぎ)は種子先端に形成される突起状の構造物で、野生イネでは自然状況下において種子の拡散や種子の保護に役立っている。
● イネはアジアとアフリカの2地域で独立に栽培化されたが、どちらの地域のイネでも芒は栽培化の過程で失う方向に選抜された。
● アジアイネとアフリカイネでは異なる遺伝子が選抜されることにより、芒喪失という共通の表現型を達成することができた。
概要
人類はおよそ1万年かけて、野生イネを改良して栽培に適したものにしてきました。イネはアジアとアフリカの2地域で独立に栽培化されましたが、その標的となった表現型は両者で共通するものが多く、芒の喪失もその1つでした。東北大学大学院生命科学研究科の別所-上原助教ら、名古屋大学そのほかの共同研究チームは、栽培化の過程でアフリカイネが芒(のぎ、種子先端にできる突起状構造物)を失う原因となった遺伝子変異を同定しました。これまでに研究チームは、アジアイネの芒喪失にRAE1とRAE2の2つの遺伝子の機能欠損が重要であったことを示してきましたが、アフリカイネの芒喪失については詳しくわかっていませんでした。本研究では、アフリカイネにおける芒喪失はRAE3という遺伝子の機能欠損が原因であったことを示しました。これまでアジアイネとアフリカイネの栽培化関連形質は、同じ遺伝子の異なる変異が選抜されることにより達成されてきたと報告されていましたが、今回初めて、アジアイネとアフリカイネで共通の栽培化形質(芒の喪失)が異なる遺伝子変異の選抜によってもたらされたことを明らかにしました。
この研究成果は、2023年1月17日付で米国科学雑誌「Proceedings of the National Academy of Sciences」電子版に掲載されました。
詳細な説明
イネは人類の活動エネルギーの約21%を供給している重要な作物です。我々が食するイネはもともと存在していたわけではなく、約1万年かけて、野生イネから栽培化されてきました。栽培化とは、雑草のように可食部が少なく、栽培しにくい形態を持つ野生イネから、人間が扱いやすくて収量が高いイネを選び出し、農業を営む上で都合の良いイネを生み出す過程のことを指します。イネの栽培化は、アジアとアフリカの2地域で起こりました。アジアでは野生種Oryza rufipogonから栽培種O. sativa(アジアイネ)が、アフリカでは野生種O. barthiiから栽培種O. glaberrima(アフリカイネ)がそれぞれ栽培化され、現在この2種のイネが食されています。栽培化においては様々な形態的・生理的特徴が標的となってきました。例えば、直立した草型注1)、種子の脱粒のしにくさ注2)、種皮の色注3)、そして、芒の喪失などがあげられます。
芒とは種子先端に形成される突起状の構造物のことで、イネ科植物に一般的に見られる特徴です。野生イネは非常に長い芒を有していますが、栽培イネでは無芒もしくは非常に短い芒がある程度です(図1)。野生イネの芒は、自然条件下においては動物の毛皮にからまることで種子拡散に寄与し、鳥や獣による食害から種子を保護する役目があります。しかし、人間が扱う上では収穫時に皮膚を傷つけたり、播種が煩雑になったり、種子貯蔵時の「かさ」が増すなど、厄介な特徴でもあったため、イネにおいては芒を消失する方向に選抜がかかったと考えられています。
私たちはこれまでにアジアのイネにおいて、O. rufipogonからO. sativaへと栽培化される過程でRegulator of Awn Elongation 1 (RAE1)とRAE2の2つの遺伝子が機能を失うことにより芒を喪失した(もしくはとても短くなった)ことを報告してきました(Furuta et al. G3 2015; Bessho-Uehara et al. PNAS 2016)。しかし、一方でアフリカの栽培種O. glaberrimaは、芒を失っているにも関わらず、RAE1とRAE2の2つの遺伝子を機能型で保持していることがわかりました。このことから、O. glaberrimaの栽培化においてはRAE1、RAE2とは別の遺伝子が選抜されることにより芒を失ったということが示唆されていました。
そこで今回、私たちはO. sativaにO. glaberrimaを戻し交配注4)した系統を用いて、遺伝学的な解析を行い、O. glaberrimaが芒を失う原因となった遺伝子RAE3を突き止めました。RAE3遺伝子は幼穂や雄しべ特異的に高発現しており、芒を形成する時期に特異的に発現していることがわかりました。またRAE3遺伝子はタンパク質をユビキチン化する機能をもつ複合体の一つの酵素であるE3ユビキチンリガーゼをコードしていました。酵母を用いた実験から、RAE3タンパク質がE1、E2酵素と協調して基質タンパク質にユビキチンを付加する機能を持つことがわかりました注5)。基質タンパク質はユビキチンが付加されることによって、(1)26Sプロテアソーム系へ輸送され分解、(2)細胞内局在が変化、もしくは(3)タンパク質の性質変化が引き起こされると考えられます。機能型RAE3は芒を伸長させることが可能であることから、RAE3タンパク質の基質となる未同定のタンパク質は芒伸長のシグナルを抑制するようなものではないかと類推されます(図2)。
また、アフリカの野生種O. barthii、栽培種O. glaberrimaとアジアの栽培種O. sativaのRAE3遺伝子配列を比較したところ、O. glaberrima特異的に48 bpの塩基欠失が生じていました。この塩基欠失は終止コドン注6)を含んでいたため、O. glaberrimaのRAE3タンパク質はO. barthiiのRAE3タンパク質に比べて長くなっていることがわかりました。また、O. barthiiとO. sativaは機能型RAE3を保持していることが明らかとなりました。E3ユビキチンリガーゼのC末端の配列は基質認識領域だと予想されています。O. glaberrimaのRAE3遺伝子ではこの領域に大幅な変異が入ることによって基質を認識できなくなり、基質のユビキチン化ができなくなったと考えられます。さらに、染色体断片部分置換系統群注7)の中から、RAE1、RAE2、RAE3がそれぞれ機能型、非機能型の複数の組み合わせ系統を選抜し、遺伝型と芒表現型を比較しました。この結果、RAE1、RAE2は芒伸長に対してそれぞれ独立に機能すること、またRAE3は単独では芒伸長に寄与せずRAE1またはRAE2のどちらかとペアになることで芒伸長を促進することが明らかとなりました(図3)。本研究により、同じシグナル経路上にある3つの遺伝子のうち、アジアではRAE1とRAE2、アフリカではRAE3という異なる遺伝子が選抜されることによって、芒を失うという同じ表現型を独立に獲得したことが示されました。
本研究成果の意義
イネは離れた2地域で栽培化されてきたにも関わらず、2つの栽培イネはよく似た形態的特徴を示します(直立した草型、難脱粒性、白い種皮色)。驚くべきことに、これらの表現型の選抜に関わるとされてこれまでに同定された遺伝子は、アジアのイネとアフリカのイネで同じものでした注1, 2, 3)。本研究により、異なる遺伝子の選抜が芒の喪失という同一表現型の獲得の原因であったことが明らかとなり、アジアとアフリカでのイネの栽培化は独立に起きたことをより強く示す証拠となりました。
用語解説
注1) 直立した草型
草の姿形のこと。野生イネでは茎が地面にくっつくほど開ききった形をしているが、それでは茎の先端にできる穂を収穫しにくいため、栽培化の過程でまっすぐ伸びる草型が選抜された。Prostrate growth 1 (PROG1) 遺伝子が栽培化の過程で選抜され、アフリカの栽培イネO. glaberrimaではこの遺伝子を丸ごと欠失し、アジアの栽培イネO. sativaでは遺伝子内に塩基置換が入ることによって機能を喪失した。
注2) 種子の脱粒のしにくさ
野生イネは、風に吹かれた時に散らばるもしくは動物に付着させることで種子を散布しており、その場合、穂から外れやすい(脱粒しやすい)方が適応的である。しかし農業を行う上では脱粒すると収量が減ってしまうため、脱粒しにくい形質が栽培化の過程で選抜され、栽培イネでは脱粒しにくくなった。OsSH1遺伝子が栽培化の過程で選抜され、O. glaberrimaではこの遺伝子を丸ごと欠失し、O. sativaでは遺伝子中の13 bpの欠失により機能を喪失した。
注3) 種皮の色
野生イネでは種皮にアントシアニンが蓄積し、赤色を呈すものが多いが、栽培化により種皮が白いものが選抜された。Rc遺伝子が栽培化の過程で選抜され、O. glaberrimaでは遺伝子内で複数の塩基置換が起き、O. sativaではO. glaberrimaのものとは異なる塩基置換ならびに塩基欠失により機能を喪失した。
注4) 戻し交配 (Backcross)
交配して作った雑種(子孫)に対して、最初の親のうち片方を再び交配すること。本研究では、まずO. sativaとO. glaberrimaを交配して作ったF1に対してO. glaberrimaを再び交配し、BC1F1 (backcross 1 time) を作出した。その後、O. glaberrimaの戻し交配をさらに2回行ったBC3F1を自殖させたBC3F2を材料に、RAE3遺伝子を突き止めた。
注5) ユビキチンの付加
E1(活性化酵素)、E2(結合酵素)、E3(ユビキチンリガーゼ)の3つの酵素の働きにより、標的タンパク質(基質タンパク質)にユビキチンを結合させる翻訳後修飾系のこと。ほぼ全ての真核生物で保存された機構であり、ユビキチンが付加された基質タンパク質は分解されたり、機能や性質が変化したりする。
注6) 終止コドン
mRNAの連続した塩基3つを1組とした配列をコドンと呼び、コドンに対応するtRNAが1つのアミノ酸を運ぶ(例えばATGというコドンはメチオニンを規定する)。終止コドンはいずれのアミノ酸にも対応せず、タンパク質翻訳を終わらせる、すなわち、翻訳を終止させる役割を果たす。
注7) 染色体断片置換系統群
CSSL(chromosome segment substitution lines)とも呼ばれる。2つの系統を交配して、戻し交配を繰り返すことにより染色体の一部だけが片方の系統に、そして染色体の大部分がもう片方の系統に置換した系統群のこと。
参考図

図1. 野生イネ(有芒)と栽培イネ(無芒)の種子。種子の上に長く伸びる突起状の構造物が芒。
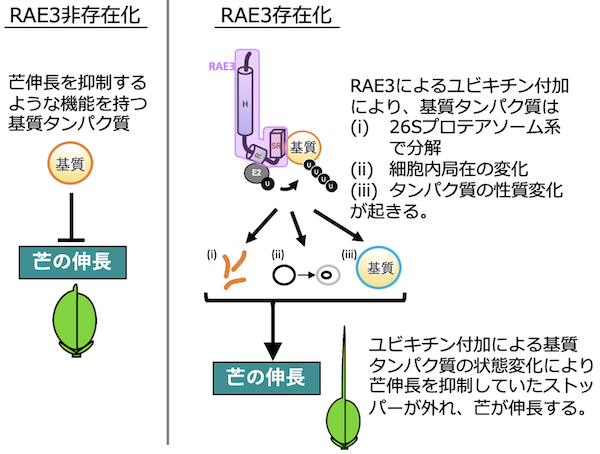
図2. 芒伸長におけるRAE3タンパク質の役割。
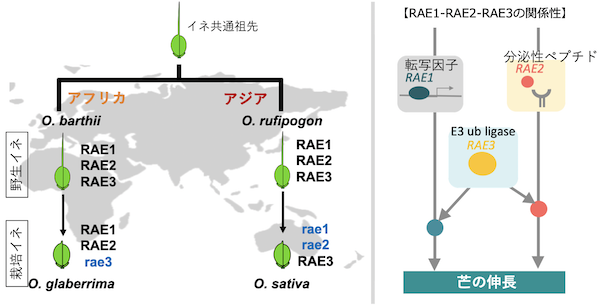
図3. 予想されるアジアとアフリカにおけるRAE1、RAE2、RAE3遺伝子の選抜過程とRAE1-RAE2-RAE3の遺伝学的関係性。
謝辞
・科研費 学術変革領域研究(A)(JP20H05912)
・科研費 研究活動スタート支援(JP20K22644)
・科研費 特別研究員奨励費(JP15J03740)
・科学技術振興機構(JST)地球規模課題対応国際科学技術協力プログラム(SATREPS)
「ミャンマーにおけるイネゲノム育種システム強化」(JPMJSA1706)
論文題目
題目:REGULATOR OF AWN ELONGATION 3, an E3 ubiquitin ligase, is responsible for loss of awns during African rice domestication
著者: Kanako Bessho-Uehara, Kengo Masuda, Diane R. Wang, Rosalyn B. Angeles-Shim, Keisuke Obara, Keisuke Nagai, Riri Murase, Shin-ichiro Aoki, Tomoyuki Furuta, Kotaro Miura, Jianzhong Wu, Yoshiyuki Yamagata, Hideshi Yasui, Michael B. Kantar, Atsushi Yoshimura, Takumi Kamura, Susan R. McCouch, Motoyuki Ashikari*
*責任著者
筆頭著者情報:(氏名、所属):別所-上原奏子(東北大学大学院生命科学研究科)
雑誌:Proceedings of the National Academy of Sciences
DOI:https://doi.org/10.1073/pnas.2207105120
著者所属
東北大学・大学院生命科学研究科(別所-上原奏子)
名古屋大学・生物機能開発利用研究センター(永井啓祐、芦苅基行)
名古屋大学・大学院生命農学研究科(増田健吾、村瀬李梨)
名古屋大学・農学部(青木振一郎)
名古屋大学・大学院生命理学研究科(小原圭介、嘉村巧)
岡山大学・資源植物科学研究所(古田智敬)
九州大学・大学院農学研究院(山形悦透、安井秀、吉村淳)
福井県立大学・生物資源学部(三浦孝太郎)
農業・食品産業技術総合研究機構(呉健忠)
パデュー大学・農学部(Diane R. Wang)
テキサス工科大学・植物土壌科学部(Rosalyn B. Angeles-Shim)
ハワイ大学・熱帯植物土壌科学部(Michael B. Kantar)
コーネル大学・植物遺伝育種学部(Susan R. McCouch)
問い合わせ先
<研究に関すること>
東北大学 大学院生命科学研究科、理学部生物学科
担当 助教 別所-上原 奏子(べっしょ-うえはら かなこ)
電話番号: 022-795-3543
Eメール: kanako.bessho.b3[at]tohoku.ac.jp
<報道に関すること>
東北大学大学院生命科学研究科 広報室
担当 高橋 さやか (たかはし さやか)
TEL:022-217-6193
Mail:lifsci-pr[at]grp.tohoku.ac.jp
*[at]を@に置き換えてください
Posted on:2023年1月19日





