
お知らせ

- トップ
- お知らせ
植物受精卵の微小管バンド移動の仕組みを解明
~エージェントモデルで読み解く過剰応答能と複数の秩序化原理~
概要
植物の茎と根を結ぶ上下軸(体軸)は,受精卵期の異方的な先端成長(1)とその後の上下不等分裂によって決まると考えられています(図1A).シロイヌナズナにおいて,受精卵の異方成長の際に,細胞の表層微小管が先端付近に円環状のバンド構造を形成し,成長に伴って細胞先端側へバンドが移動することが,実験から観察できていました(図1B).しかし,どのような仕組みで多数の微小管が協調あるいは競合して配向秩序化(2)しバンド構造を形成し,かつ移動を達成するのかは未解明でした.
本研究では,秋田県立大学の野々山朋信博士研究員および津川暁助教らと東北大学の植田美那子教授らが協働し,顕微鏡画像から微小管バンド幅とバンド移動速度を定量化しました. この定量データに基づいて,微小管エージェントモデル(3)を構築し,微小管の配向を変化させる方向誘因領域に対して微小管の応答能(4)や微小管バンド移動の必要条件を定量的に示しました.類似の先端成長を示す根毛細胞などでは成長が速すぎるためにバンドが形成されない可能性が示唆されました.
これらの知見により,表層微小管に代表される細胞骨格と細胞形態の関係が明らかになるばかりでなく,方向誘引により繊維バンド構造が集積するような繊維強化材料などの設計や製造への応用も期待されます.
発表のポイント
1. シロイヌナズナの受精卵が異方成長する際に形成される微小管バンド構造とその移動現象の仕組みを,数値シミュレーションによって定量的に明らかにしました.
2. 微小管の配向を変化させる方向誘因領域よりも微小管バンドが幅広く形成されることや,微小管がバンドを形成する際に異なる2つの仕組みがあり得ることを示しました.
3. これらの数値シミュレーション結果は,微繊維の配向形成やその移動を制御できる可能性を示しており,繊維強化材料の設計などへの応用が期待されます.
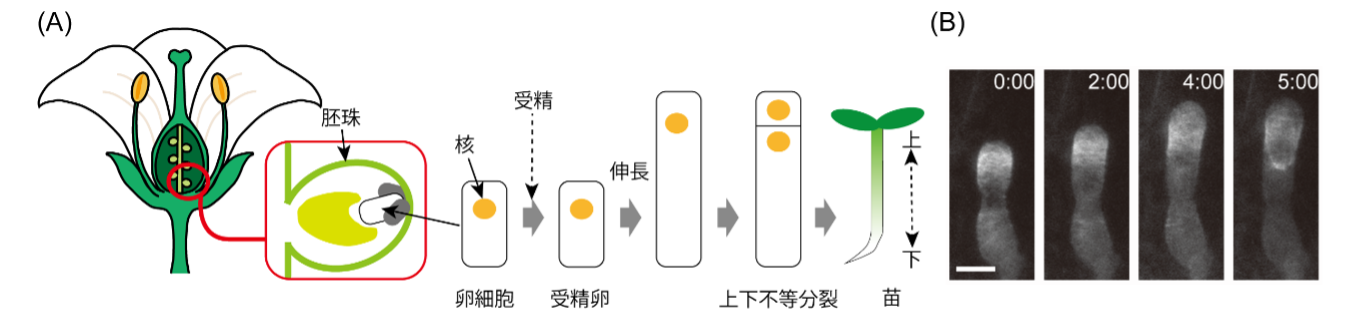
図1:受精卵の異方成長期に形成される微小管バンド構造とその移動現象
(A)被子植物(シロイヌナズナ)における受精卵は一方向的に異方的に伸長することで個体の上下軸を作る.胚珠の中にある受精卵が異方成長し上下不等分裂することで個体の上下軸が決まると考えられている.(B)顕微鏡によるシロイヌナズナの受精卵の伸長を撮影した画像.時間経過とともに微小管が円環バンド構造を形成し,細胞先端側へ移動する(白).
成果掲載誌
論文タイトル:Agent-based Simulation of Cortical Microtubule Band Movement in Arabidopsis Zygotes (シロイヌナズナ受精卵の表層微小管バンド移動現象のエージェントシミュレーション)
著者:Kanako Bessho-Uehara*, Tomoki Omori, Stefan Reuscher, Keisuke Nagai, Ayumi Agata, Mikiko Kojima, Tomonobu Nonoyama,Zichen Kang,Hikari Matsumoto,Sakumi Nakagawa,Minako Ueda,Satoru Tsugawa
雑誌名:Scientific Reports
DOI:10.1038/s41598-025-11078-8
詳細な説明
研究の背景
植物には細胞形態を維持し細胞内外の運動に必要な物理的力を発生させる細胞内の繊維状構造が存在し,細胞骨格と呼ばれています.東北大学の植田美那子教授らのグループによって,植物受精卵の表層微小管が円環状に整列してバンド構造が形成されることが発見されていましたが,そのバンド形成の仕組みは詳しくわかっていませんでした.また,受精卵を24時間程度観察すると,微小管バンドが細胞成長に伴って細胞先端側へ移動することもわかってきましたが,ナノメートルスケールの微小管がどのように配向秩序化し整列するかは長く謎に包まれていました.そこで本研究では,微小管のイメージング画像分析と,微小管をエージェントと見立てた手法であるエージェントモデルを組み合わせることで,分子スケールの振る舞いから細胞スケールのバンド移動現象の仕組みを解明することを目指しました.
本研究の成果
本研究では,まず2光子顕微鏡を用いた蛍光観察を基に,受精卵内の微小管バンドの幅と移動速度を定量化しました(図2A).その結果,微小管バンドは細胞先端付近に維持され,細胞成長速度とほぼ同じ速度で先端側へ移動していることが明らかになりました(図2B).この実験データを分子レベルの振る舞いから説明するために,表層微小管の分子生物学的挙動を考慮したエージェントモデルを構築しました(図2C).この手法では,微小管のプラス端が0.08μm/sで伸長するという実験事実や,微小管同士が衝突する際に束になる現象や,ある条件下では微小管が短縮する現象などが12個の主要なパラメータによって組み込まれています.これらのパラメータ設定によって,受精卵内部の表層で微小管がどのように振る舞うかを分析することができます.私たちはこのモデルを使って,微小管バンド形成には微小管が伸長する方向を変化させる方向誘因が必要であると考え,方向誘因の強さを段階的に変えた場合にいつどのように微小管バンドができるかを調べました.
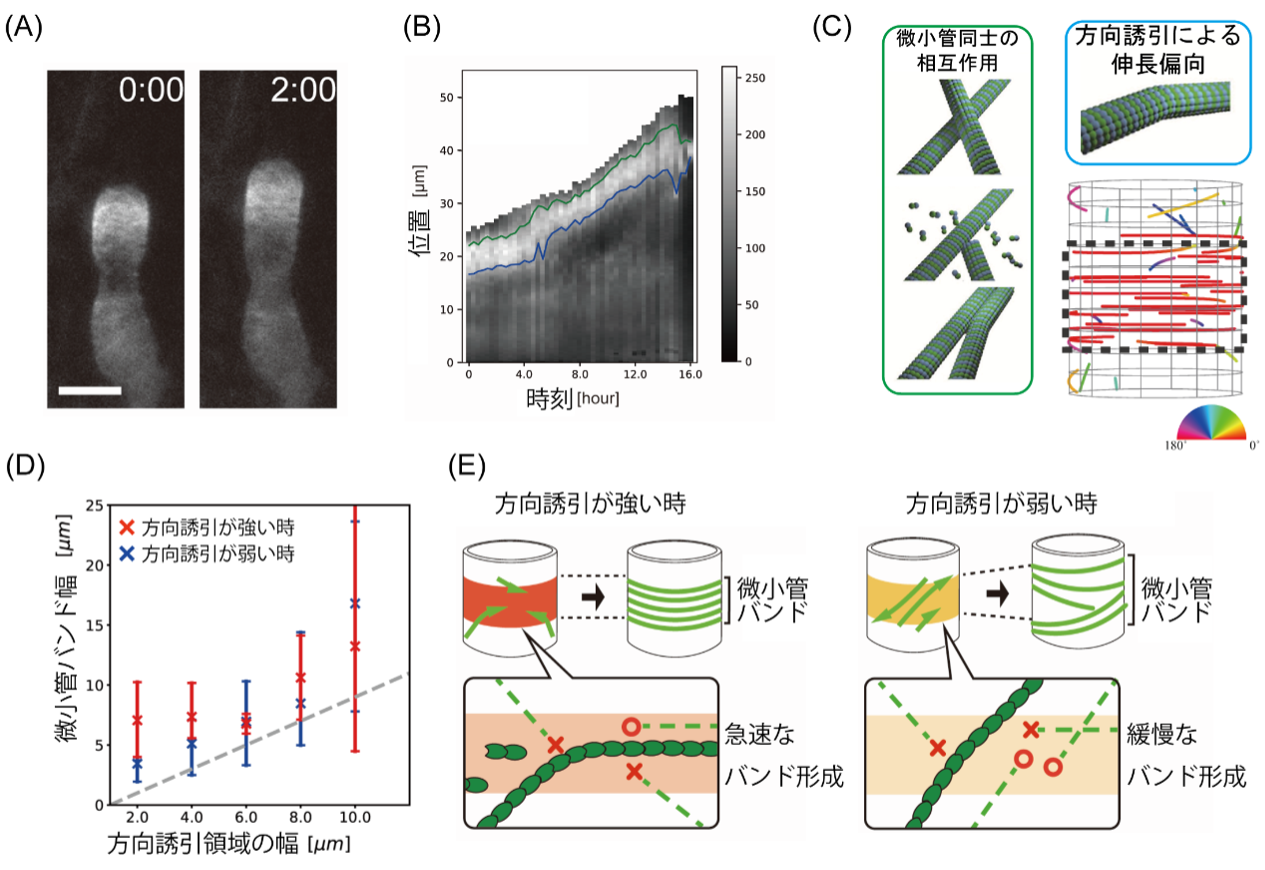
図2:微小管バンドの過剰応答能を示すシミュレーション結果図.
(A)顕微鏡による受精卵の表層微小管を撮影した画像. 白色部が表層微小管の局在を示す. (B)時間経過とともに微小管バンドが先端側へ移動する様子(上限を緑線,下限を青線で示す).(C)微小管同士の相互作用と方向誘引領域(黒点線領域)を導入した微小管エージェントモデルの概念図. カラープロットは微小管の配向角度を表す.(D)入力する方向誘引領域の幅と出力される微小管バンド幅の関係.(E)領域内の方向誘引の強弱によって, 急速にバンドが形成される場合と緩慢にバンドが形成される場合が存在することを発見した.
その結果,方向誘因の強さに依らず,方向誘因領域よりも幅広い領域に微小管バンドが形成される,という非自明な結果を得ました(図2D).また,方向誘因の強さによって,微小管バンドが急激に整列する場合と,緩慢に整列する場合の異なる2つの仕組みがあるもわかりました(図2E).これらの結果は驚くべき結果で,入力としての微小管方向の制御と実際の微小管集団運動が異なることを意味しています.
これらの微小管バンドの幅やバンド構造のでき方に関する入力と出力の差異に加えて,方向誘因領域を細胞先端側に移動させる場合にも興味深い結果を得ました.方向誘因領域の速度を実験データに近い入力速度に設定した場合には微小管バンドが形成されるのに対し, 実験データ範囲外の高速な入力速度の場合には微小管バンドが方向誘因領域に追いつけず取り残されることがわかりました.この結果は,類似の先端成長細胞である受精卵よりも速い細胞成長速度を持つ根毛や花粉管などで微小管バンドが形成されないことの要因である可能性が示唆されました.
今後の期待
植物受精卵内で起こる生理学的な現象を知るためには顕微鏡観察が有効ですが,細胞スケールよりもさらに小さいナノスケールの微小管の精密な観察には限界があります. 本研究のように,分子レベルの実験データを取り込んだエージェントモデルは,観察が難しい分子の振る舞いを推測するために有効であり,細胞スケールと分子スケールを横断的につなぐ重要な手法であると考えられます.このような集団運動シミュレーションは,受精卵などの発生生物学における分子の振る舞いを説明するだけでなく,微細な繊維を制御することで製造される繊維強化材料や複合材料などへの応用も期待されます.
用語解説
(1)先端成長:細胞の先端表面のみが伸展する成長様式.植物の根毛や花粉管などでよく見られる.
(2)配向秩序化:数多くの分子の向き(配向)が時間経過とともに同じ向きに揃うことを指す.
(3)エージェントモデル:多数の因子やエージェントがあるルールによって動きを変えたときに全体の振る舞いを調べる計算手法.例えば,道路網において多数の車が動き,複数の車が目的地までたどり着く時間を調べる方法などがある.
(4)微小管の応答能:微小管が外部刺激に対して伸長速度や向きを変える度合いを指す.
研究体制と支援
本研究は, 秋田県立大学(野々山朋信博士研究員,津川暁 助教),東北大学(植田美那子教授ら)の共同研究として行われました.
本研究は, 文部科学省の科学研究費補助金 (JP20K15832, JP22K15135, JP25K18499, JP19H05670, JP19H05676, JP23H02494, JP22K21352, JP25KJ0540),科学技術振興機構(JST)の戦略的創造研究推進事業(JPMJCR2121),公益財団法人サントリー生命科学財団(SunRiSE)と公益財団法人東レ科学振興会(20-6102)の支援を受けて行われました.
問い合わせ先
<研究に関すること>
東北大学大学院生命科学研究科、兼担 理学部生物学科[web]
教授 植田美那子
TEL: 022-795-6713
Email: minako.ueda.e7[at]tohoku.ac.jp
<報道に関すること>
東北大学大学院生命科学研究科広報室
高橋 さやか(たかはし さやか)
TEL:022-217-6193
Email:lifsci-pr[at]grp.tohoku.ac.jp
*[at]を@に置き換えてください
Posted on:2025年7月30日





